- Дафнии
-

В данной статье или разделе имеется список источников или внешних ссылок, но источники отдельных утверждений остаются неясными из-за отсутствия сносок. Вы можете улучшить статью, внеся более точные указания на источники.Дафния 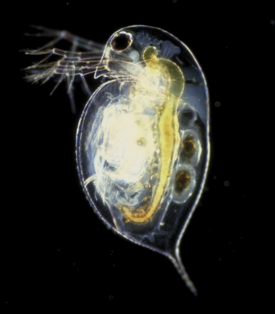 Daphnia pulex (Leydig, 1860)
Daphnia pulex (Leydig, 1860)Научная классификация Царство: Животные Тип: Членистоногие Надкласс: Ракообразные Класс: Жаброногие Надотряд: Ветвистоусые Отряд: Daphniiformes Подотряд: Anomopoda Семейство: Daphniidae Род: Дафния Латинское название Daphnia O. F. Müller, 1785 Виды См. в тексте статьи.
ITIS 83873 NCBI 6668
Дафнии (лат. Daphnia) — род планктонных ракообразных из надотряда ветвистоусых (Cladocera). Размерами от 0,2 до 6 мм в длину. Иногда дафний называют водяными блохами[1].Содержание
Систематика
Один из наиболее крупных (более 50 валидных видов в мировой фауне) и сложных для систематики родов ветвистоусых. Типовой вид — D. longispina O.F. Mueller, 1785. Наиболее характерный отличительный признак рода — сросшиеся с головой антенны I самок. Кроме того, у самок обычно хорошо развит рострум, а вентральный край створок выпуклый. У обоих полов створки, как правило, несут шипики и образуют непарный вырост — хвостовую иглу. У большинства видов (кроме некоторых австралийских видов, часто относимых к роду Daphniopsis) в эфиппиуме два яйца. Все щетинки антенн II обычные, с длинными сетулами.
Большинство систематиков признают деление этого рода на два подрода — Daphnia (Daphnia) O.F. Mueller, 1785 и Daphnia (Ctenodaphnia) Dybowski et Grochjwski, 1895. У представителей подрода Daphnia (Daphnia) отсутствует выемка головного щита, яйцевые камеры эфиппиума обычно почти перпендикулярны спинному краю створок. У представителей подрода Daphnia (Ctenodaphnia) выемка головного щита имеется, яйцевые камеры эфиппиума обычно почти параллельны спинному краю створок. Все виды, ранее относимые к роду Daphniopsis), входят в состав подрода Daphnia (Ctenodaphnia), причем многие из них обладают примитивными (отсутствие выемки головного щита) или уклоняющимися (одно яйцо в эфиппиуме) признаками.
Виды дафний
- Daphnia ambigua Scourfield, 1947
- Daphnia atkinsoni Baird, 1859
- Daphnia arcuata Forbes, 1893
- Daphnia australis (Sergeev et Williams, 1985)
- Daphnia barbata Weltner, 1898
- Daphnia bolivari Richard, 1888
- Daphnia brooksi Dodson, 1985
- Daphnia carinata King, 1852
- Daphnia carvicervix Ekman, 1901
- Daphnia catawba Coker, 1926
- Daphnia cheraphila Hebert and Finston, 1996
- Daphnia chevreuxi Richard, 1896
- Daphnia cristata G. O. Sars, 1862 — дафния хохлатая
- Daphnia cucullata G. O. Sars, 1862)
- Daphnia curvirostris Eylmann, 1887
- Daphnia dadayana Daday, 1902
sensu Paggi, 1999 - Daphnia dubia Herrick, 1883
- Daphnia dolichocephala G.O. Sars, 1895
- Daphnia ephemeralis (Schwartz et Hebert, 1985)
- Daphnia exilis Herrick, 1895
- Daphnia galeata G. O. Sars, 1864
- Daphnia gessneri Herbst, 1968
- Daphnia hispanica Glagolev et Alonso, 1990
- Daphnia hyalina Leydig, 1860
- Daphnia jollyi Petkovski, 1973
- Daphnia lacustris G.O. Sars, 1862
- Daphnia laevis Birge, 1879
- Daphnia latispina Korinek and Hebert, 1996
- Daphnia longiremis G. O. Sars, 1862
- Daphnia longispina O. F. Mueller, 1785)
- Daphnia lumholtzi G. O. Sars, 1885
- Daphnia magna Straus, 1820
- Daphnia magniceps Herrick, 1884
- Daphnia mediterranea Alonso, 1985
- Daphnia menucoensis Paggi, 1996
- Daphnia middendorffiana Fischer, 1851
- Daphnia minnehaha Herrick, 1884
- Daphnia nivalis Hebert, 1977
- Daphnia occidentalis Benzie, 1986
- Daphnia obtusa Kurz, 1875
- Daphnia oregonensis Korinek and Hebert, 1996
- Daphnia pamirensis Rylov, 1928
- Daphnia parvula Fordyce, 1901
- Daphnia peruviana Harding, 1955
- Daphnia pileata Hebert and Finston, 1996
- Daphnia prolata Hebert and Finston, 1996
- Daphnia psittacea Baird, 1850
- Daphnia pulex Leydig, 1860)
- Daphnia pulicaria Forbes, 1893
- Daphnia pusilla (Serventy, 1929)
- Daphnia retrocurva Forbes, 1882
- Daphnia quadrangula (Sergeev, 1990)
- Daphnia queenslandensis (Sergeev, 1990)
- Daphnia rosea G. O. Sars, 1862
- Daphnia salina Hebert and Finston, 1993
- Daphnia schoedleri G. O. Sars, 1862
- Daphnia similis Claus, 1876
- Daphnia similoides Hudec, 1991
- Daphnia sinevi Kotov, Ishida et Taylor, 2006
- Daphnia stuederi (Ruhe, 1914)
- Daphnia tanakai Ishida, Kotov et Taylor, 2006
- Daphnia tenebrosa G. O. Sars, 1898
- Daphnia tibetana (G.O. Sars, 1903)
- Daphnia thomsoni G.O. Sars, 1894
- Daphnia thorata Forbes, 1893
- Daphnia triquetra G.O. Sars, 1903
- Daphnia truncata Hebert et Wilson, 2000
- Daphnia turbinata G.O.Sars, 1903
- Daphnia umbra
- Daphnia villosa Korinek and Hebert, 1996
- Daphnia wardi Hebert et Wilson, 2000
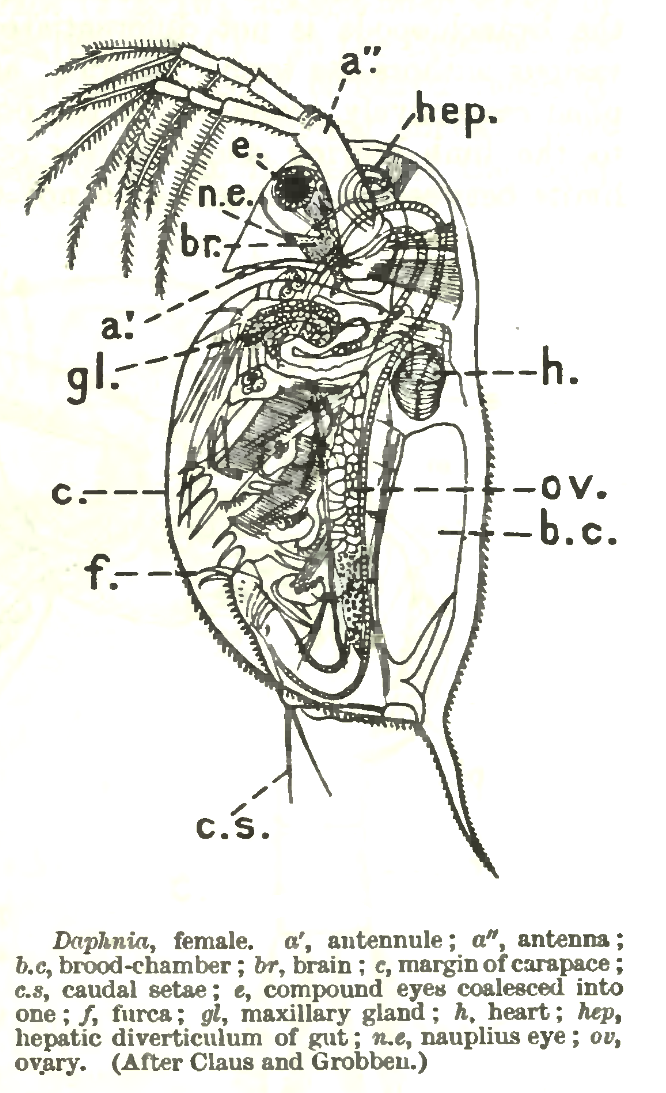
Внешнее строение
Если не сказано иначе, в этом разделе описано строение самок. Покровы состоят из головного щита и двустворчатого карапакса. Обычно на них имеется хорошо заметный узор из ромбов и многоугольников — ретикуляция. Каждую такую ячейку покровов образует одна клетка гиподермы. На краю створок имеются шипики, а на заднем конце — покрытая шипиками хвостовая игла. У многих видов на внутреннем крае створок в его средней части есть ряд оперенных щетинок, у самцов всех видов есть такие же щетинки и добавочные щетинки на передне-нижнем углу створок.
На голове у большинства видов развит клювовидный вырост — рострум. Под ним расположены первые антенны (антеннулы) — короткие выросты, несущие на конце 9 обонятельных щетинок — эстетасок, а на боковой поверхности — одну добавочную щетинку. У самцов антенны первые более крупные, подвижные, несут кроме эстетасков крупную щетинку («жгутик») на дистальном конце.
На боковой поверхности головы расположены выступы кутикулы — форниксы. Их форма, а также форма заднего края головного щита — важный диагностический признак подродов и групп видов. Под форниксами к голове сложным «суставом» крепятся вторые антенны (антенны). Они состоят из основания и двух ветвей — внутренней трёхчлениковой и наружной четырёхчлениковой. На концах члеников ветвей расположены покрытые уплощёнными волосками двучленистые плавательные щетинки, образующие «весла» при плавании. На трёхчлениковой ветви их пять (четыре только у D. cristata), на четырёхчлениковой — четыре. На основании есть несколько мелких чувствительных щетинок.
От задней поверхности головы отходит крупная верхняя губа. Внутри неё находятся несколько гигантских полиплоидных клеток, которые выделяют секрет, склеивающий пищу в пищевой комок.
На границе между головным щитом и карапаксом под створками расположены мандибулы. Они имеют сложную форму, асимметричны и имеют сильно хитинизированные жевательные поверхности, покрытые гребнями и выростами. В процессе питания мандибулы переправляют пищу в ротовое отверстие.
Под карапаксом расположены небольшие максиллы первые (максиллулы), несущие четыре щетинки. Максиллы вторые у дафний редуцированы. Имеется пять пар двуветвистых грудных ног сложного строения. Ноги первой и отчасти второй пары различаются по строению у самцов и самок. На первой паре ног самцов расположены крючковидные выросты, которые позволяют цепляться к самкам при спаривании. Третья и четвёртая пары несут веера фильтрующих щетинок. На каждой ноге имеется дыхательный придаток — эпиподит.
За грудным отделом расположен редуцированный брюшной, наличие которого «маркируется» дорзальными абдоминальными выростами, закрывающими выход из выводковой камеры. Их обычно четыре, они хорошо развиты у половозрелых самок и редуцированы у самцов большинства видов.
Задний отдел тела — крупный подвижный постабдомен, гомологичный тельсону других ракообразных. На его дорзальной стороне расположены два ряда зубчиков, между которыми находится анальное отверстие. У самцов некоторых видов эти зубчики частично или полностью редуцированы. На конце постабдомена имеются парные коготки, покрытые шипиками. По одним данным они гомологичны фурке, по другим — представляют собой пару крупных видоизмененных щетинок. Шипики есть на наружной и внутренней стороне коготков, обычно на наружной стороне три группы шипиков, на внутренней — две. Постабдомен служит для очистки фильтрационного аппарата от крупных посторонних частиц.
Внутреннее строение
 Схема строения дафнии
Схема строения дафнии
 Голова дафнии
Голова дафнииПокровная система дафний представлена типичной гиподермой. Гиподерма карпакса состоит из крупных клеток, образующих ячейки ромбической формы.
Центральная нервная система состоит из надглоточного ганглия (головного мозга) и брюшной нервной цепочкой с несколькими парными ганглиями. Головной мозг хорошо заметен у живых особей, что встречается крайне редко. Он состоит из крупного, разделенного пополам оптического ганглия и собственно надглоточного ганглия. От передней части оптического ганглия отходит оптический нерв, соединяющий мозг со сложным фасеточным глазом. Непарный сложный глаз у дафний образуется из парного зачатка (у эмбрионов два глаза) и содержит ровно 22 фасетки (омматидия). Он расположен в особой полости внутри головы, к стенкам которой подвешен с помощью двух связок (лигаментов) и приводится в движение тремя парами глазодвигательных мышц. У живых особей заметен тремор глаза, время от времени наблюдаются более крупные скачки (саккады) глаза. От головного мозга отходят нервы также к глазку (простому глазу), первым антеннам (в их основании имеется чувствительный ганглий, клетки которого иннервируют обонятельные щетинки — эстетаски), а также нервы к чувствительному затылочному органу неизвестного назначения. К нижней части надглоточного ганглия примыкает простой глаз (глазок, науплиальный глаз). У большинства видов он содержит пигмент и заметен в виде небольшой чёрной точки. Вокруг пигментного пятна расположены 4 группы чувствительных клеток.
У дафний имеется сложная система поперечнополосатых мышц, приводящих в движение вторые антенны, постабдомен и грудные конечности, а также мышцы, приводящие в движение глаз, верхнюю губу и др. Мышцы пищеварительного тракта также поперечнополосатые.
Пищеварительный тракт начинается ротовым отверстием, которое прикрыто крупной верхней губой. Гигантские высокополиплоидные клетки, расположенные внутри губы, выделяют секрет, склеивающий пищу в пищевой комок. Движениями мандибул он переправляется в тонкий пищевод, мышцы-дилататоры которого создают перистальтику, обеспечивая транспорт пищи по пищеводу. Внутри головы пищевод переходит в более широкую среднюю кишку, которая тянется до средней части постабдомена. Внутри головы от средней кишки отходят два изогнутых печеночных выроста. В задней части постабдомена находится короткая задняя кишка.
Сердце находится на спинной стороне тела, спереди от края выводковой камеры. Кровь (гемолимфа), ток которой хорошо заметен благодаря наличию в ней бесцветных клеток — фагоцитов, попадает внутрь сердца через остии — два щелевидных боковых отверстия. При сокращении сердца остии закрываются клапанами, а кровь выбрасывается через переднее отверстие к голове. Кровеносные сосуды отсутствуют, закономерное направление токов крови обеспечивается прозрачными перегородками между разными участками миксоцеля.
Дыхание происходит через покровы тела, в первую очередь грудных ног, на которых имеются дыхательные придатки — эпиподиты. Последние принимают участие также в осморегуляции. Дополнительный орган осморегуляции новорожденных особей — крупная затылочная пора (затылочный орган), который исчезает после первой постэмбриональной линьки.
Органы выделения — сложной формы максиллярные железы, которые расположены на внутренней поверхности створок в их передней части.
Парные яичники (у самцов — семенники) расположены по бокам от кишечника. На заднем конце находится зона размножения оогониев, остальная часть яичника заполнена созревающими ооцитами. Яйцеклетки по мере созревания перемещаются к их задней трети, где расположены узкие яйцеводы, открывающиеся в выводковую камеру. У самцов семепроводы открываются на постабдомене в его дистальной части, у многих видов на специальных сосочках.
Линька
При линьке расходится цервикальный шов — линия между головным щитом и карапаксом, и животное вылезает из экзувия. Вместе с карапаксом сбрасываются покровы тела и конечностей. Линька периодически повторяется в течение всей жизни особи. Обычно линька происходит в толще воды, эфиппиальные самки некоторых видов линяют, приклеившись снизу к поверхностной пленке воды. Несколько линек происходят во время эмбрионального развития, в выводковой камере.
Распространение
Род Daphnia имеет всесветное распространение (включая Антарктиду, где в реликтовых солёных озёрах оазиса Вестфолль (Vestfold Hills) была обнаружена Daphnia studeri, ранее относимая к роду Daphniopsis). В начале XX века преобладало мнение о космополитном распространении большинства видов, однако впоследствии выяснилось, что фауны разных континентов сильно различаются. Некоторые виды, тем не менее, имеют очень широкие ареалы и распространены на нескольких материках. Наименьшее число видов характерно для экваториальных районов, где дафнии редки. Наиболее разнообразна фауна субтропиков и умеренных широт. В послединие десятилетия ареалы многих видов изменились из-за их расселения человеком. Так, в Европу (Англию) был вселен вид из Нового Света D. ambigua. Во многих водохранилищах юга США стала обычной D. lumholtzi, которая до этого встречалась только в Старом Свете.
В прудах и лужах средней полосы России часто встречаются (и наиболее популярны среди аквариумистов) следующие рачки рода дафния:
Дафния магна (D. magna), самка — до 6 мм, самец — до 2 мм, новорожденные — 0,7 мм. Созревают в течение 10—14 суток. Пометы через 12—14 суток. В кладке до 80 яиц (обычно 20—30). Продолжительность жизни — до 3-х месяцев.
Дафния пулекс (D. pulex), самка — до 3—4 мм, самец — 1—2 мм. Пометы через 3—5 суток. В кладке до 25 яиц (обычно 10—12). Живут 26—47 суток.
В озёрах умеренной зоны Евразии часто встречаются D. cucullata, D. galeata, D. cristata и несколько других видов.Биология
Дафнии — мелкие рачки (размеры тела взрослых особей от 0,6 до 6 мм). Они населяют все типы стоячих континентальных водоемов, встречаются также во многих реках с медленным течением. В лужах, прудах и озёрах часто имеют высокую численность и биомассу. Дафнии — типичные планктонные рачки, большую часть времени проводящие в толще воды. Различные виды населяют мелкие временные водоемы. литораль и пелагиаль озёр. Довольно многие виды, особенно населяющие засушливые районы — галофилы, обитающие в солоноватых, соленых и гипергалинных континентальных водоемах. К таким видам относятся, например, D. magna, D. atkinsoni, D.mediterranea, a также большинство видов, ранее относимых к роду Daphniopsis.
Локомоция
Большую часть времени проводят в толще воды, передвигаясь резкими скачками за счет взмахов вторых антенн, которые покрыты специальными оперенными щетинками (отсюда их обычное название — «водяные блохи», часто относимое ко всем ветвистоусым). Многие дафнии способны также медленно ползать по дну или стенкам сосудов за счет токов воды, создаваемых грудными ножками (антенны при этом способе передвижения неподвижны).
Питание
Основной пищей для дафний служат бактерии и одноклеточные водоросли. Виды, которые переживают зиму в активном состоянии (в глубоких, непромерзающих водоемах), проводят её в придонных слоях воды, питаясь в основном детритом. Питаются путем фильтрации, создавая токи воды ритмическими движениями грудных ножек. Пища отфильтровывается веерами фильтрующих щетинок, которые расположены на эндоподитах III и IV пар грудных ног. Крупные частицы, застрявшие в фильтрационном аппарате (например, нитчатые водоросли), удаляются с помощью постабдомена и его коготков. С фильтрующих вееров пища поступает в брюшной пищевой желобок, передается к максиллам первой пары и затем к мандибулам, движения которых переправляют её в пищевод[2]. Спереди ротовое отверстие дафний прикрыто крупной верхней губой, внутри которой расположены слюнные железы из гигантских полиплоидных клеток. Их секрет склеивает частицы пищи в пищевой комок.
При средней концентрации корма в воде, взрослые дафнии различных видов фильтруют со скоростью от 1 до 10 мл/сутки. Суточное потребление пищи взрослой D. magna может достигать 600 % от веса её тела[3].
Газообмен
На грудных ножках расположены мешковидные дыхательные придатки — жабры. Вероятно, значительную часть кислорода дафнии получают через тонкие покровы тела и конечностей, а дыхательные придатки, как и затылочный орган новорожденных, играют важную роль в осморегуляции. Некоторые виды (например, D. pulex, D. magna) при пониженном содержании кислорода в воде начинают синтезировать гемоглобин, так что их гемолимфа и всё тело окрашивается в красный цвет.
Цикломорфоз
Многие виды дафний (в основном обитающие в озёрах) обладают цикломорфозом — разные их поколения, развивающиеся в различные сезоны года, резко различаются по форме тела. В умеренных широтах летние поколения таких видов имеют удлиненные выросты кутикулы — хвостовую иглу и шлем головного щита. У весенних и осенних поколений хвостовая игла короче, шлем может быть короче или вообще отсутствует. Показано, что отращивание выростов требует энергетических затрат и приводит к снижению плодовитости. Из факторов, вызывающих рост выростов покровов, было продемонстрировано влияние повышенной турбулентности воды, высокой температуры и др. Позднее было показано, что основное влияние на рост цикломорфных выростов оказывают кайромоны — сигнальные вещества, выделяемые разными видами беспозвоночных и позвоночных хищников. Выдвигалось множество гипотез об адаптивной роли цикломорфоза: облегчение парения в менее плотной и вязкой воде, ускоренное плавание в горизонтальном направлении и др. Большинство из них либо не доказаны, либо опровергнуты. Сейчас в качестве основного объяснения цикломорфоза принимается теория о роли выростов в защите от хищников. Прозрачные выросты увеличивают реальный размер тела и защищают тем самым от мелких беспозвоночных хищников — более крупную добычу с выростами труднее схватить и не выпустить при манипулировании — «запихивании в рот». Иногда при этом хвостовая игла обламывается, что может играть ту же роль, что и автотомия хвоста у ящерицы. В то же время прозрачные выросты не увеличивают видимого размера, что важно для защиты от крупных зрительных хищников — рыб.
Вертикальные миграции
Как и другие представители зоопланктона, дафнии совершают вертикальные миграции во многих водоемах — как крупных и глубоких, так и мелких. Практически всегда эти миграции носят суточный характер: в светлое время суток рачки перемещаются в более глубокие придонные слои, а в темное время суток — поднимаются к поверхности. Среди множества теорий, объясняющих причины миграций, на которые затрачивается заметный процент энергии, наиболее обоснованной сейчас считается теория, связывающая миграции всё с той же защитой от хищников. В светлое время суток более крупные виды, хорошо заметные для зрительных хищников, вынуждены опускаться в глубокие, плохо освещенные слои воды для защиты от выедания рыбами. Миграции у одного и того же вида дафний могут быть хорошо выражены в озёрах, где рыб много, и отсутствовать в озёрах, где планктоядные рыбы отсутствуют.
Размножение
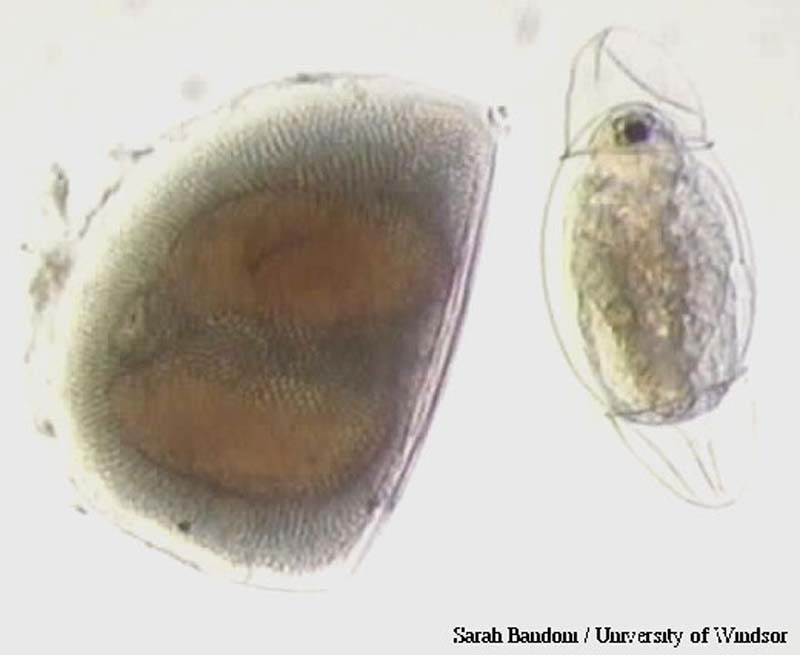
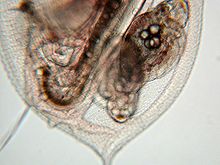 Выводковая камера дафнии с эмбрионами
Выводковая камера дафнии с эмбрионами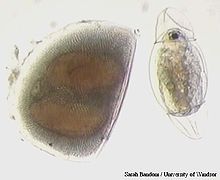 Эфиппиум Daphnia (Daphnia) и новорожденная самка, только что вылупившаяся из покоящегося яйца
Эфиппиум Daphnia (Daphnia) и новорожденная самка, только что вылупившаяся из покоящегося яйцаДля большинства дафний (как и других ветвистоусых) характерен циклический партеногенез. В благоприятных условиях (например летом в непересыхающих лужах и мелких водоёмах) в популяциях дафний присутствуют только партеногенетические самки. Из их неоплодотворенных диплоидных яиц развиваются следующие поколения самок. Поскольку партеногенез у дафний амейотический, их размножение клональное (популяции состоят из клонов — потомства отдельных самок). Развитие эмбрионов при партеногенезе протекает в выводковой камере под раковинкой и сопровождается несколькими линьками. Затем молодые рачки выходят из выводковой камеры (развитие прямое).
Осенью или перед высыханием водоема из таких же неоплодотворенных яиц развиваются самцы (как правило, все особи одного помета имеют одинаковый пол). Таким образом, определение пола у дафний чисто средовое.
Переход к двуполому размножению у видов из крупных водоемов обычно требует воздействия двух стимулов — понижения температуры и уменьшения длины светового дня. Предполагается, что эти и другие стимулы действуют через снижение потребления пищи самкой. Самцы дафний имеют более мелкие размеры, существенно отличаются от самок по строению первых антенн и грудных ног. Они плавают быстрее самок и спариваются с ними, прикрепившись к заднему краю раковинки. Оплодотворение у дафний внутреннее.
В последние годы показано, что развитие самцов может быть индуцировано добавлением в среду ювенильного гормона ракообразных метил-фарнезоата, а также аналогов ювенильного гормона насекомых — инсектицида феноксикарба[4].
В период двуполого размножения одни самки рождают самцов, а другие в это же время образуют покоящиеся, или эфиппиальные, яйца. Они образуются путем мейоза и для развития требуют оплодотворения. После оплодотворения они также выходят в выводковую камеру, покровы которой утолщаются и образуют специальную хитиновую камеру — эфиппиум (эфиппий). Развитие яиц продолжается до стадии гаструлы, затем они впадают в анабиоз. При очередной линьке самка сбрасывает эфиппиум, который у некоторых видов обычно опускается на дно, а у других плавает на поверхности водоема. При откладке эффипия самки некоторых видов (например, D. pulex) нередко погибают. Вместе с эфиппиумами яйца дафний разносятся ветром, распространяются на лапах млекопитающих, на лапах и перьях птиц, а также в их кишечниках. Яйца в химически стойкой оболочке внутри эфиппия после прохождения через кишечник птиц и рыб могут оставаться жизнеспособными. Обычно они также выдерживают промерзание и длительное высыхание. Показано, что покоящиеся яйца дафний могут длительное время выживать и развиваться в растворах ядовитых солей (например, хлорида ртути HgCl2) при концентрации ядов, в тысячи раз превышающей ПДК[5]; после разрыва оболочки яйца вылупившиеся эмбрионы в таких растворах сразу же погибают.
Использование в научных исследованиях
Дафнии используются в качестве модельных организмов во многих экологических, токсикологических и генетических исследованиях.
Расшифровка генома
За последние годы осуществлена частичная расшифровка генома дафнии Daphnia pulex, в 2011 г. был завершен его черновик[6]. Геном дафнии состоит из 200 миллионов нуклеотидов, но при этом содержит минимум 30,9 тысяч генов — больше, чем у других до сих пор изученных многоклеточных животных (например в геноме человека около 20-25 тысяч генов). Геному дафнии оказался свойственен высокий темп генных дупликаций, что привело к созданию многочисленных генных кластеров. Более трети продуктов генов, обнаруженных в геноме дафнии, не имеют известных гомологов в протеомах других организмов. Наиболее крупные амплифицированные генные семейства также характерны только для этой эволюционной линии. Многие гены-паралоги экспрессируются по-разному в зависимости от условий среды; уникальные для дафний гены наиболее чувствительны к изменениям условий. Можно предполагать, что генные дупликации увеличили экологическую пластичность дафний, позволяя им приспособиться к условиям разных водоёмов и к меняющимся условиям в одном водоёме[7][8][9].
Разведение
Широко распространено культивирование дафний в качестве кормового объекта, как в промышленных целях[10], так и любителями — аквариумистами. В благоприятных условиях дафнии легко и быстро размножаются и растут, что позволяет получать 30—50 (в отдельных случаях — до 100) граммов рачков в сутки с одного кубического метра культуры[11].
Исходная культура легко может быть получена из природных водоёмов. Желательно отлавливать рачков в небольших водоёмах, где популяция дафний может быть практически свободна от примеси других животных. В зимнее время культуру дафний можно получить из эфиппиальных покоящихся яиц, заранее собранных с поверхности воды или из верхнего слоя ила. Собранные эфиппиумы хорошо сохраняются в сухом состоянии в прохладном помещении[12].
В зависимости от необходимого количества рачков, культивировать дафний можно как в небольших сосудах, так и в крупных бассейнах и прудах. Оптимальная плотность культуры — 300—1000 г/м³[12]. Периодически, раз в несколько недель или месяцев, культуру перезакладывают. Старение культуры связано с накоплением в ней продуктов обмена и разложения и с засорением её другими организмами. При подмене воды срок жизни культуры продлевается.
Оптимальная температура выращивания — 15—25°С, реакция среды нейтральная (pH 6,8—7,8), содержание кислорода — не менее 3—6 мг/л, окисляемость 14,8—26,2 мг O2/л[13].
При выращивании дафний применяют и совместное, и раздельное культивирование рачков и корма для них.
При совместном выращивании в культуру добавляют органические удобрения, например, навоз, вносимый в количестве 1,5 кг/м3. Возможно выращивание на минеральных удобрениях, добавка которых вызывает бурное развитие одноклеточных водорослей.
Недостатком совместного культивирования является сильное загрязнение воды, быстрое старение культуры и быстрое зарастание емкости нитчатыми водорослями.
Раздельное выращивание дафний и корма для них лишено этих недостатков, но технически сложнее и применяется в основном в условиях массового промышленного культивирования кормовых рачков. При этом отдельно выращивается водорослевая культура, которая 1—2 раза в день вносится в ёмкость с дафниями.
В лабораторных и домашних условиях удобно содержать культуру дафний на дрожжах, вносимых ежедневно понемногу в количестве 15—20 г на кубический метр культуры (15—20 мг/л). [14]. Стандартизированные методы лабораторного культивирования дафний описаны в руководствах по токсикологии и биотестированию[15].
Прочие факты
Живые, сушеные и замороженные дафнии нередко используются как корм для аквариумных рыбок или насекомых, содержимых в террариумах. В промышленном рыбоводстве большое значение имеет разведение дафний на корм.
Дафнии — один из стандартных объектов для тестирования токсичности водных растворов химических соединений, применяемых в исследовании загрязнений водной среды[16]. Дафнии чувствительны даже к небольшим концентрациям некоторых солей, например, добавление солей меди в концентрации 0,01 мг/л вызывает замедление движений рачков, они либо опускаются на дно, либо замирают у поверхностной плёнки воды[17].
Примечания
- ↑ Дафния // Энциклопедический словарь Брокгауза и Ефрона: В 86 томах (82 т. и 4 доп.). — СПб., 1890—1907.
- ↑ Ивлева, 1969, с. 119
- ↑ Ивлева, 1969, с. 123
- ↑ Keonho Kim, Alexey A Kotov, and Derek J. Taylor. Hormonal induction of undescribed males resolves cryptic species of cladocerans//Proceedings of the Royal Society B: Biological Sciences. 2006 January 22; 273(1583): 141—147.
- ↑ Макрушин, 2006
- ↑ Colbourne et al, 2011, с. 555
- ↑ Daphnia Genome Database
- ↑ Геном рачка-дафнии содержит больше генов, чем геном человека
- ↑ Найдено животное с самым большим количеством генов — Lenta.ru
- ↑ Ивлева, 1969, с. 128
- ↑ Ивлева, 1969, с. 136
- ↑ 1 2 Ивлева, 1969, с. 129
- ↑ Шпет, 1950
- ↑ Ивлева, 1969, с. 134
- ↑ Руководство…, 2002, с. 48
- ↑ Руководство…, 2002
- ↑ Ивлева, с. 1щ3
Литература
- Peters P.H., De Bernardi R. Daphnia // Mem. Ist. Ital. Idrobiol. — 1987. — V. 45. — 502 p.
- John K. Colbourne, Michael E. Pfrender, Donald Gilbert, W. Kelley Thomas, Abraham Tucker, Todd H. Oakley, Shinichi Tokishita, Andrea Aerts, Georg J. Arnold, Malay Kumar Basu, Darren J. Bauer, Carla E. Cáceres, Liran Carme, Claudio Casola, Jeong-Hyeon Choi, John C. Detter, Qunfeng Dong, Serge Dusheyko, Brian D. Eads, Thomas Fröhlich, Kerry A. Geiler-Samerotte, Daniel Gerlach, Phil Hatcher, Sanjuro Jogdeo, Jeroen Krijgsveld1, Evgenia V. Kriventseva, Dietmar Kültz, Christian Laforsch, Erika Lindquist, Jacqueline Lopez, J. Robert Manak, Jean Muller, Jasmyn Pangilinan, Rupali P. Patwardhan, Samuel Pitluck, Ellen J. Pritham, Andreas Rechtsteiner, Mina Rho, Igor B. Rogozin, Onur Sakarya, Asaf Salamov, Sarah Schaack, Harris Shapiro, Yasuhiro Shiga, Courtney Skalitzky, Zachary Smith, Alexander Souvorov, Way Sung, Zuojian Tang, Dai Tsuchiya, Hank Tu, Harmjan Vos, Mei Wang, Yuri I. Wolf, Hideo Yamagata, Takuji Yamada1, Yuzhen Ye, Joseph R. Shaw, Justen Andrews, Teresa J. Crease, Haixu Tang, Susan M. Lucas, Hugh M. Robertson, Peer Bork, Eugene V. Koonin, Evgeny M. Zdobnov, Igor V. Grigoriev, Michael Lynch, Jeffrey L. Boore. The Ecoresponsive Genome of Daphnia pulex // Science. — 2011. — Т. 331. — № 6017. — С. 555—561. — DOI:10.1126/science.1197761
- Ивлева И. В. Биологические основы и методы массового культивирования кормовых беспозвоночных. — М.: «Наука», 1969.
- Макрушин А. В., Лянгузова И. В. Оболочка пропагул беспозвоночных и растений: избирательная проницаемость и барьерные свойства // Журнал общей биологии. — 2006. — Т. 67. — № 2. — С. 120—126.
- Шпет Г. И. Разведение дафний как живого корма в рыбоводстве // Труды Украинского ин-та пруд. и озёрно-речн. рыбн. хоз-ва. — 1950. — Т. 7.
- Руководство по определению методом биотестирования токсичности вод, донных отложений, загрязняющих веществ и буровых растворов. — М.: РЭФИА, НИА-Природа, 2002.
Ссылки
- Daphnia Genome Database — база данных о геномах представителей рода Daphnia (англ.) (Проверено 5 марта 2011)
Категории:- Животные по алфавиту
- Пресноводные ракообразные
- Жаброногие
- Аквакультура
- Аквариумный корм




Wikimedia Foundation. 2010.